
The Biofilm Matrix - A Living Engineering Material
Chenxi Zhai, Haoyuan Shi, Tianjiao Li, Liming Zhao, Jingjie Yeo
J2 Lab for Engineering Living Materials, Sibley School of Mechanical and Aerospace Engineering, Cornell University
Introduction
Engineering materials are increasingly equipped with living organisms to imbue autonomous biological characteristics, such as specific and targeted recognition of environmental changes, self-repair, and self-assembly. Despite current fruitful integrations with biomaterials, such as cell-embedded stimuli-responsive materials [1,2], these living components usually passively react to ambient changes. The autonomy of the living organisms is not fully brought to bear. Therefore, Engineered Living Materials (ELMs) are an emerging area of research aimed at empowering materials with greater intelligence manifesting as autonomous growth, independent judgment, and active regulation of their material properties. For instance, autonomous growth will allow the ELMs absorb energy and matter from their natural environments to construct structures using biologically-producible materials, in contrast to requiring specifically synthesized components to form these structures [3]. These forms of material autonomy may be achieved by genetic engineering and spatial engineering [4,5].
In particular, bacteria are promising organisms that can be incorporated into engineering materials due to rapid developments in synthetic biology, such as bacterial genetic modification and tracking technology. Crucially, bacteria can secrete extracellular polymeric substances (EPS) with potentially desirable engineering features when forming biofilms, in terms of their unique and emergent structural, mechanical, and biochemical properties. In the biofilm matrix, the EPS form biopolymeric hydrogels that typically consist of extracellular DNA, polysaccharides, lipids, and proteins. The EPS is produced after bacteria adhere to a substrate. Planktonic bacterial cells recognize substrates via their flagella [6] and sense the membrane deformation during attachment [7]. Once bacteria start to colonize the substrate, these planktonic cells transit to sessile phenotypes [8,9], accompanied by a series of physiological regulations that help bacteria survive and settle on the substrate.
An important part of this strategy is the secretion of EPS. sEPS, a type of micromolecules in the EPS, mediates the surface properties of substrates, such as the hydrophilicity, to facilitate the initial adhesion as well as to promote subsequent cell deposition on the first layer of bacteria [9,10]. Other components in the EPS provide bridges for connecting neighboring bacteria cells to accumulate clusters and to link those clusters into a larger matrix [9,11]. Mature biofilm matrices have cross-linked spatial structures and mechanical properties that are comparable with synthetic polymeric gels [12], with the additional potential benefits of autonomous growth, repair, and stimuli-response of the biofilm matrix. By naturally encapsulating living organisms, the biofilm matrix can be manipulated by engineering the constitutive bacteria and biofilms using synthetic biology. Additionally, bacterial secretion of EPS is more energy-efficient than artificial chemical synthesis of polymeric substrates [13]. Therefore, the biofilm matrix is anticipated to be a vital material pillar in developing the next generation of intelligent engineering structures and materials.
To facilitate deeper understanding and broader discussions on this “biofilm-as-a-material” paradigm, we introduce the biofilm matrix from the following perspectives: (1) the multiscale structures, (2) the mechanical properties, and (3) applications for engineering living materials.
Multiscale Structures
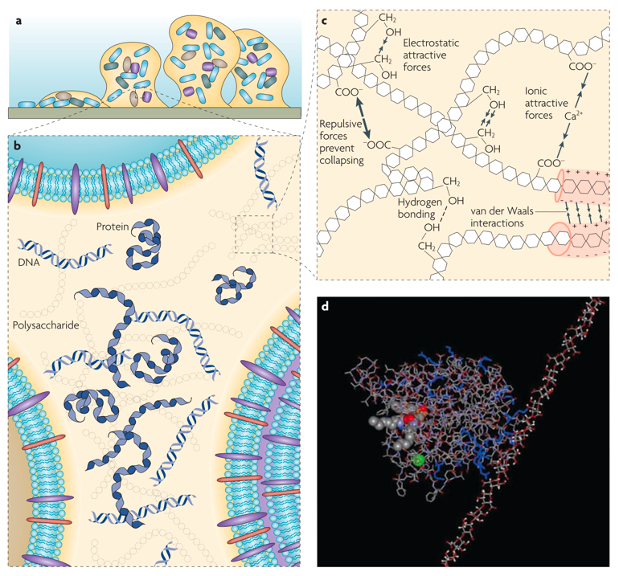
Figure 1. (a) A model of a bacterial biofilm attached to a solid surface. (b) Polysaccharides, proteins and DNA are distributed between the cells in a non-homogeneous pattern, setting up differences between regions of the matrix (c) The classes of weak physicochemical interactions and the entanglement of biopolymers that dominate the stability of the EPS matrix (d) A molecular modelling simulation of the interaction between the exopolysaccharide alginate (right) and the extracellular enzyme lipase (left) of Pseudomonas aeruginosa in aqueous solution. Adapted from [14].
The EPS in the biofilm matrix play two major roles in a biofilm system. First, the biopolymers aggregate and crosslink to form a three-dimensional scaffold encasing the living organisms. Second, the EPS ensure the mechanical stability of biofilms, provide adhesion to surfaces, and render cohesion in the biofilm [14]. EPS are composed of a large variety of biopolymers, so large that their multiscale heterogenous structures and properties are difficult to characterize thoroughly, thereby earning them the monikers of “the dark matter of biofilms” or “the great unexplored” [9,14]. For different biofilm systems, the exact composition of the EPS varies depends on the species of the constitutive microorganisms and the mechanical, thermal, and biochemical conditions of their environment. Generally, EPS are composed of extracellular polysaccharides (exopolysaccharides) and proteins, along with nucleic acids [15]. Collectively, these components form multiscale structures ranging from molecular aggregates to macroscale polymer networks (Figure 1).
Exopolysaccharides are polymers with high molecular weights (~ 20 to 500 kDa) while being linear and/or branched, and are composed of diverse monosaccharides [14]. These biopolymers are secreted by most species of bacteria, and they are usually the most abundant component in a biofilm matrix [3]. Within the matrix, networks of crosslinked exopolysaccharides provide the architecture in which the cells are aggregated, immobilized, and protected, hence they are also referred to as the “molecular glue” [3]. These networks can be visualized using electron microscopy [16]. However, from the perspective of engineering living materials, structurally modifying the polysaccharides via genetic engineering is inherently difficult due to the lack of standardization in biosynthetic pathways across species [3].
Extracellular proteins are generally in the forms of enzyme and structural proteins. The enzymes empower the ecological system of EPS by sequestering nutrients from the environment and degrading the EPS components during starvation. The structural, non-enzymatic proteins are crucial for forming coherent biofilms and stabilizing the exopolysaccharide networks. These proteins also link the bacteria’s cellular surface to the EPS networks [14]. For example, amyloid proteins facilitate the adhesion of biofilms to the surface. The persistence length of these amyloid fibrils can range from tens of nanometers to the scale of microns [17]. The mechanical properties of such natural semiflexible fibrils can be significantly superior to that of synthetic fibers [18], with reported Young’s moduli in the range of 0.2 to 20 GPa [17,19,20]. As these proteins are directly encoded genetically, it is relatively easier to engineer protein structures and functions, in contrast to the difficulties encountered with exopolysaccharides. This relative ease makes these proteins attractive candidates for modifying and equipping biofilms with specific functionalities [3].
Extracellular DNA (eDNA) is found in biofilms of various origins [14]. The basis of eDNA formation is not simply due to cell lysis. In fact, evidence shows that cells can actively excrete DNA molecules into the EPS [21]. eDNA can be one of the major matrix components in biofilms, such as those of Pseudomonas aeruginosa, by functioning as an intercellular connector and stabilizing the biofilm [22]. eDNA also serves as an adhesin in Bacillus cereus biofilms [23]. eDNA assemblies mostly occur on the sub-cellular scale. Even though eDNA assemblies are extensively observed and explored experimentally, there is a dearth of good methods for engineering the eDNA to induce super-cellular structures for the biofilms.
Mechanical Properties
Understanding the mechanical performance of biofilms is important for designing strategies to properly tackle biofilm-related problems [24] as well as for conceiving bacteria-based ELMs. The physical response of biofilms to their ambient environment is determined by their mechanical properties. However, it is still challenging to fully grasp the mechanical details of biofilms through experiments and computational modeling due to their dynamically-responsive nature. Here, we will elucidate the general mechanical properties of biofilms.
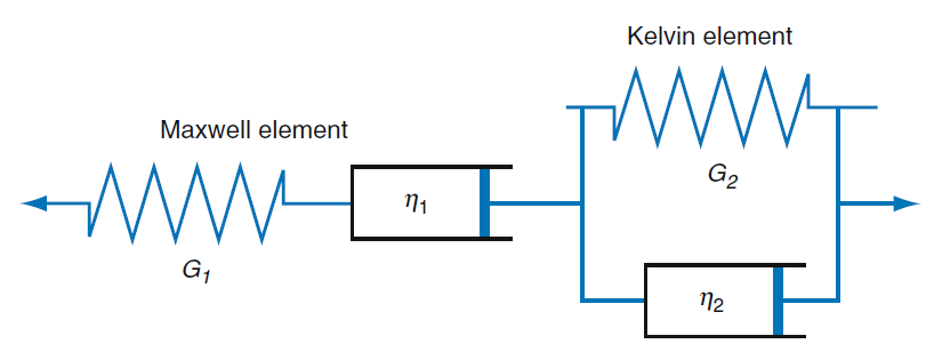
Stress relaxation and cone rheometer are two important methods for characterizing the viscoelastic properties of biofilms. More specifically, stress relaxation of biofilms can be determined using low load compression testing (LLCT) experimentally [25] and finite element methods computationally. Streptococcus mutans biofilms displayed viscoelastic characteristics in microrheological studies, being elastic in short-range (within the elastic limit) while plastic in long-range (beyond the elastic limit). However, Cense et al. found that biofilms had viscoelastic solid characteristics using stress-relaxation tensile tests [26]. This difference vividly demonstrates the structural heterogeneity at different scales of the biofilms that can lead to a broad variety of mechanical properties, where the EPS play an important role [26]. In addition, biofilm stiffness was higher during growth while being softer during starvation [27]. A plasticity model was used to describe the plastic and failure behavior of biofilms, and to calculate the tensile strength [28]. This study showed the irreversible plastic deformation in biofilms. Biofilm’s viscoelastic behavior and response was also modelled with a Burger’s model of springs and dashpots (Figure 2) [24]. The springs were used to model the reversible elastic behavior and the dashpots were used to model irreversible plastic behavior, which corresponded well with the experiments. In general, finite element models can be used to match the numerical displacement fields with those from experiments to determine more accurate mechanical properties [24].
Figure 2. Burger’s model of springs and dashpots to simulate viscoelastic behavior in biofilms, reproduced from reference [24].
Rheologically, shear forces from fluids can passively remove cells from the biofilm matrix, which negatively impacts the stability of the biofilm matrix. For instance, shear stresses resulted in the phenotypic adaptation of the bacteria [29]. However, polysaccharide synthesis locus (PSL) cross-linking in the EPS could form stable structures and improved the viscoelasticity buffering and resisting of rapidly changing shear forces [30,31]. The biofilm behaved as a viscoelastic solid when the shear stress was below the critical biofilm growing stress and as a viscoelastic fluid when the shear stress was above it. The shear modulus of the biofilm was found to increase in tandem with increasing shear stresses [29,32]. The deformation of biofilms by fluid flow and interactions between phases was systematically studied with a novel thermodynamic model [31], by which the biofilm cohesive failure was accurately predicted. In general, biofilm viscoelastic properties vary significantly with shear stresses, biofilm growing conditions, and bacterial strain. Also, the precise mechanisms dictating biofilm surface morphology by structural and regulatory components need further elucidation. Mechanical instabilities, such as wrinkling and delamination, were found to affect the growth morphology of biofilms. Interfacial energy was found to be the key driving force of mechanomorphogenesis through theoretical simulations and mechanical measurements [33].
Adhesive strength is defined as the work required to separate the 29-day-old grown P. fluorescens biofilms from their attachment [34]. Biofilm adhesive and cohesive strengths are important indicators of biofilm failure since adhesive failure will lead to the separation of the biofilm from the surface [35]. The adhesive strength of biofilms was varies with fluid velocity, age, glucose concentration, pH, and roughness [36]. For example, when the glucose concentration was above 30 mg/L, the adhesive strength would decrease as the high concentration interfered with biofilm adhesion. Several components in the EPS contribute to the strong adhesive properties of biofilms, such that strongly adhering materials can be synthesized using these components. For instance, bacterial curli fibers were used to prepare strong adhesives and DeBenedictis et al. used atomistic molecular dynamics (MD) simulations to examine curli subunit adsorption on surfaces. The adhesive property was found to be significantly influenced by both structural features and sequence influence [37]. Furthermore, adhesive behavior and detachment mechanisms of curli nanofibers were elucidated by coarse-grained (CG) MD simulations, revealing three different peeling process [38].
Applications
Self-healing Concrete
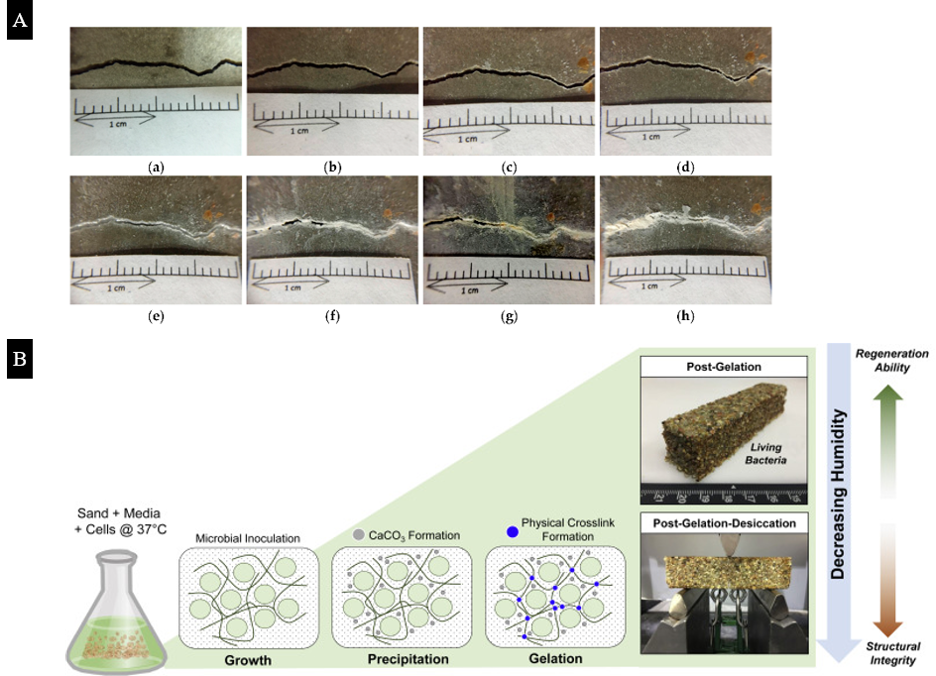
The biological activity of microorganisms embedded in materials can induce nanoscale changes within the system in response to external stimuli and effect macroscale alterations. These complex responses are mediated by the ability of the microorganisms to survive for extended periods of time, implying that successful establishment of biofilm matrices will be critical issues that need be considered as developments in ELMs accelerate. For example, self-healing concrete [39–42] exploits calcium carbonate (CaCO3) precipitation induced by bacteria to repair concrete cracks (Figure 3A). When the cracks are exposed, CaCO3 precipitates and adheres to the crack surface to complete a healing process. Microorganisms alter the composition of microenvironments by their metabolic activity and produce CO32-, which then forms CaCO3 biominerals in an environment that is rich in Ca2+ ions. Thus, microbially induced calcium carbonate precipitation strongly depends on external conditions, such as the pH value, humidity, and temperature [42], but the metabolic activity of microorganisms varies for different bacteria [41]. Although ureolytic bacteria is most commonly used in inducing calcium carbonate precipitation, the selection of microorganisms should be based on the pertinent environment that is relevant for the intended applications.
Figure 3. (A) The repair process of pre-cracked concrete beam specimens with large cracks (Experimental group II): (a) Day 1; (b) Day 3; (c) Day 7; (d) Day 14; (e) Day 21; (f) Day 28; (g) Day 56; (h) Day 91. (B) Formation of the LBM Mineralized Scaffold. Adapted from Reference [40,43].
Recently, Heveran et al. created living building materials (LBMs) that “grow”, where an inert scaffold of sand and hydrogel support the living organisms [43] (Figure 3B). The materials can be regulated by temperature and humidity switches for microbial-induced CaCO3 precipitation. Compared with abiotically controlled materials, LBMs exhibit higher fracture energy after desiccating, which shows the potential of designing various bio-functional materials by leveraging microorganisms. However, the limitations for bacterial self-healing concrete are also distinct, mainly due to the inhibition of bacterial vitality at high pH values and unsuitable humidity conditions [43,44]. In addition to utilizing encapsulation techniques and other porous carriers for bacteria [41], methods for maintaining the long-term viability of microorganisms need to be explored further. Also, it is necessary to consider the influence of microbial metabolic byproducts on the self-healing process and the recovery of the mechanical properties of the structure. It is likely that these issues may be circumvented by manipulating the multiscale structure, mechanics, and biochemistry of the biofilm matrix to enhance the protection and proliferation of the microbial population.
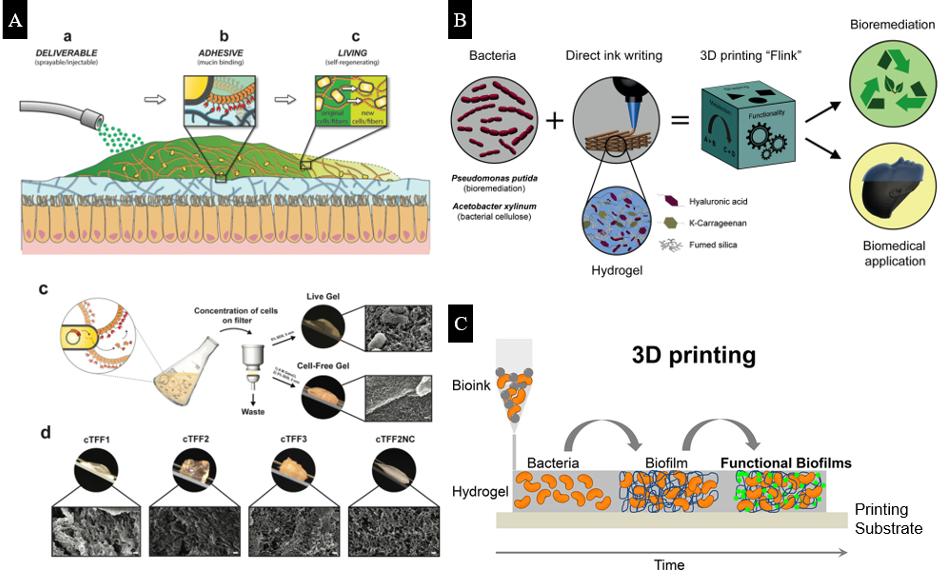
Figure 4. (A) Delivery, adhesion, and self‐regeneration of live mucoadhesive hydrogels in the human gut, and bio-fabrication of curli‐based hydrogels by engineered bacteria. (B) Schematics of the 3D bacteria-printing platform for the creation of functional living materials. (C) 3D printing of bacteria via alginate chemistry. Adapted from Reference [45–47].
Bacterial Hydrogels
A hydrogel matrix can mimic the EPS produced in a biofilm matrix, thereby acting as an artificial bacteria-hydrogel biofilm [48,49]. In addition to simulating the characteristics of actual biofilms, bacterial hydrogels have strong potential for applications as biomedical and pharmaceutical materials. Kandemir et al. studied the interactions between bacterial cells and hydrogels, showing that changes in the extracellular polymer matrix lead to changes in cell structure and metabolic functions [49]. Mainardi et al. used ionotropic gelation and alumina nanoparticles to form a porous 3D network and then embedded the bacterial species of E. coli and B. subtilis [50]. Their results demonstrate long-term bacterial viability, which means that the nanocomposites are suitable for encapsulating matrix for bacteria.
The Live Gel, designed by Joshi et al. (Figure 4), exhibits the ability to grow and self-renew. This gel has prospective biomedical applications in physical wound therapies inside the gut lumen [45]. It contains E. coli as its cellular chassis with engineered curli nanofibers as a scaffold. Compared with other adhesive materials that aim to improve the efficiency of delivering encapsulated drugs, the regenerative ability of the Live Gel allows the gel to extend its residence times in the gastrointestinal tract. Furthermore, genetic programmability may be used to dynamically alter the adhesion properties of the gel for binding with specific tissue surfaces.
Combined with 3D printing technology, bacterial hydrogels can be synthesized with specific geometries and functionalities. Schaffner et al. developed a functional living ink, “Flink,” which contains 1:1:1 ratio of hyaluronic acid (HA), ĸ-carrageenan (ĸ-CA), and fumed silica (FS). “Flink” possesses viscoelastic properties that are required for 3D printing (Figure 4B) [46]. Two types of bacterial hydrogels with P. putida and A. xylinum are printed, and these gels are able to degrade pollutants and produce bacterial cellulose, respectively. Meyer et al. developed a low-cost 3D printer for bacterial printing via alginate chemistry, where the bacterial suspension in the alginate solution can solidify on a calcium-containing substrate (Figure 4C) [51]. The 3D printed, engineered E. coli biofilm remains intact and stable in the hydrogel, which provides an ideal platform for fabricating biofilm-derived products [47,51].
Conclusions
To facilitate deeper understanding and broader discussions on the “biofilm-as-a-material” paradigm, we introduced the biofilm matrix from the following perspectives: (1) the multiscale structures, (2) the mechanical properties, and (3) applications for engineering living materials. ELMs are an emerging area of research aimed at empowering engineering materials with greater intelligence manifesting as autonomous growth, independent judgment, and active regulation of their material properties. Bacteria are particularly promising candidates for incorporation into ELMs, resulting from the incredible pace of advances in synthetic biology for manipulating the myriad biological properties of bacteria. An important part of this “biofilm-as-a-material” paradigm is bacteria’s ability to secrete EPS that confer unique and emergent structural, mechanical, and biochemical properties to the biofilm matrix that are robust and dynamically-resonsive to changes in the environment or other external stimuli. However, this multiscale complexity and heterogeneity that are responsible for the hardy behavior of the biofilm can also complicate precise characterization with existing theoretical, computational, and experimental approaches. Therefore, as the development and applications of ELMs broaden, we anticipate that a “Materials 4.0” approach will be needed to rationally design ELMs by intimately and harmoniously integrating theoretical mechanics, big data in biological omics, computational modelling and simulations, artificial intelligence (AI), and experimental approaches. With this post, we hope to spark further interdisciplinary conversations and seed future collaborations within the iMechanica community and beyond.
References
[1] X. Chen, L. Mahadevan, A. Driks, O. Sahin, Bacillus spores as building blocks for stimuli-responsive materials and nanogenerators, Nat. Nanotechnol. 9 (2014) 137–141. doi:10.1038/nnano.2013.290.
[2] C.M. Wells, M. Harris, L. Choi, V.P. Murali, F.D. Guerra, J.A. Jennings, Stimuli-responsive drug release from smart polymers, J. Funct. Biomater. 10 (2019). doi:10.3390/jfb10030034.
[3] P.Q. Nguyen, N.-M.D. Courchesne, A. Duraj-Thatte, P. Praveschotinunt, N.S. Joshi, Engineered Living Materials: Prospects and Challenges for Using Biological Systems to Direct the Assembly of Smart Materials, Adv. Mater. 30 (2018) 1704847. doi:10.1002/adma.201704847.
[4] J. Huang, S. Liu, C. Zhang, X. Wang, J. Pu, F. Ba, S. Xue, H. Ye, T. Zhao, K. Li, Y. Wang, J. Zhang, L. Wang, C. Fan, T.K. Lu, C. Zhong, Programmable and printable Bacillus subtilis biofilms as engineered living materials, Nat. Chem. Biol. 15 (2019) 34–41. doi:10.1038/s41589-018-0169-2.
[5] M. Florea, H. Hagemann, G. Santosa, J. Abbott, C.N. Micklem, X. Spencer-Milnes, L. De Arroyo Garcia, D. Paschou, C. Lazenbatt, D. Kong, H. Chughtai, K. Jensen, P.S. Freemont, R. Kitney, B. Reeve, T. Ellis, Engineering control of bacterial cellulose production using a genetic toolkit and a new celluloseproducing strain, Proc. Natl. Acad. Sci. U. S. A. 113 (2016) E3431–E3440. doi:10.1073/pnas.1522985113.
[6] R. Belas, of Surfaces By Bacteria, Trends Microbiol. 22 (2014) 517–527. doi:10.1016/j.tim.2014.05.002.
[7] H.J. Busscher, H.C. Van Der Mei, G. Subbiahdoss, P.C. Jutte, J.J.A.M. Van Den Dungen, S.A.J. Zaat, M.J. Schultz, D.W. Grainger, Biomaterial-associated infection: Locating the finish line in the race for the surface, Sci. Transl. Med. 4 (2012). doi:10.1126/scitranslmed.3004528.
[8] P. Stoodley, K. Sauer, D.G. Davies, J.W. Costerton, Biofilms as complex differentiated communities, Annu. Rev. Microbiol. 56 (2002) 187–209. doi:10.1146/annurev.micro.56.012302.160705.
[9] J. Flemming, Hans-Curt , Neu, Thomas R., Wingender, The perfect slime, 2016. doi:10.2166/9781780407425.
[10] B. Tansel, D.Z. Tansel, Adhesion strength and spreading characteristics of EPS on membrane surfaces during lateral and central growth, Colloids Surfaces B Biointerfaces. 111 (2013) 594–599. doi:10.1016/j.colsurfb.2013.07.005.
[11] C. Pellicer-Nàcher, C. Domingo-Félez, A.G. Mutlu, B.F. Smets, Critical assessment of extracellular polymeric substances extraction methods from mixed culture biomass, Water Res. 47 (2013) 5564–5574. doi:10.1016/j.watres.2013.06.026.
[12] C. Even, C. Marlière, J.M. Ghigo, J.M. Allain, A. Marcellan, E. Raspaud, Recent advances in studying single bacteria and biofilm mechanics, Adv. Colloid Interface Sci. 247 (2017) 573–588. doi:10.1016/j.cis.2017.07.026.
[13] A. Maitra, K.A. Dill, Bacterial growth laws reflect the evolutionary importance of energy efficiency, Proc. Natl. Acad. Sci. 112 (2015) 406 LP – 411. doi:10.1073/pnas.1421138111.
[14] H.C. Flemming, J. Wingender, The biofilm matrix, Nat. Rev. Microbiol. 8 (2010) 623–633. doi:10.1038/nrmicro2415.
[15] J.N.C. Fong, F.H. Yildiz, Biofilm matrix proteins, Microb. Biofilms. (2015) 201–222.
[16] J.B. Rollefson, C.S. Stephen, M. Tien, D.R. Bond, Identification of an extracellular polysaccharide network essential for cytochrome anchoring and biofilm formation in Geobacter sulfurreducens, J. Bacteriol. 193 (2011) 1023–1033.
[17] T.P. Knowles, A.W. Fitzpatrick, S. Meehan, H.R. Mott, M. Vendruscolo, C.M. Dobson, M.E. Welland, Role of intermolecular forces in defining material properties of protein nanofibrils, Science (80-. ). 318 (2007) 1900–1903.
[18] Y. Zhang, E.P. DeBenedictis, S. Keten, Cohesive and adhesive properties of crosslinked semiflexible biopolymer networks, Soft Matter. 15 (2019) 3807–3816.
[19] A.W.P. Fitzpatrick, S.T. Park, A.H. Zewail, Exceptional rigidity and biomechanics of amyloid revealed by 4D electron microscopy, Proc. Natl. Acad. Sci. 110 (2013) 10976–10981.
[20] T.P.J. Knowles, M.J. Buehler, Nanomechanics of functional and pathological amyloid materials, Nat. Nanotechnol. 6 (2011) 469–479.
[21] U. Böckelmann, A. Janke, R. Kuhn, T.R. Neu, J. Wecke, J.R. Lawrence, U. Szewzyk, Bacterial extracellular DNA forming a defined network-like structure, FEMS Microbiol. Lett. 262 (2006) 31–38.
[22] L. Yang, K.B. Barken, M.E. Skindersoe, A.B. Christensen, M. Givskov, T. Tolker-Nielsen, Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa, Microbiology. 153 (2007) 1318–1328.
[23] S. Vilain, J.M. Pretorius, J. Theron, V.S. Brözel, DNA as an adhesin: Bacillus cereus requires extracellular DNA to form biofilms, Appl. Environ. Microbiol. 75 (2009) 2861–2868.
[24] T. Guélon, J.-D. Mathias, P. Stoodley, Advances in Biofilm Mechanics, in: Springer Ser. Biofilms, Springer, Berlin, Heidelberg, 2011: pp. 111–139. doi:10.1007/978-3-642-19940-0_6.
[25] B.W. Peterson, H.J. Busscher, P.K. Sharma, H.C. Van Der Mei, Visualization of microbiological processes underlying stress relaxation in pseudomonas aeruginosa biofilms, in: Microsc. Microanal., Cambridge University Press, 2014: pp. 912–915. doi:10.1017/S1431927614000361.
[26] A.W. Cense, E.A.G. Peeters, B. Gottenbos, F.P.T. Baaijens, A.M. Nuijs, M.E.H. van Dongen, Mechanical properties and failure of Streptococcus mutans biofilms, studied using a microindentation device, J. Microbiol. Methods. 67 (2006) 463–472. doi:10.1016/j.mimet.2006.04.023.
[27] S.S. Rogers, C. Van Der Walle, T.A. Waigh, Microrheology of bacterial biofilms in Vitro: Staphylococcus aureus and pseudomonas aeruginosa, Langmuir. 24 (2008) 13549–13555. doi:10.1021/la802442d.
[28] J.L. Chaboche, A review of some plasticity and viscoplasticity constitutive theories, Int. J. Plast. 24 (2008) 1642–1693. doi:10.1016/j.ijplas.2008.03.009.
[29] P. Stoodley, R. Cargo, C.J. Rupp, S. Wilson, I. Klapper, Biofilm material properties as related to shear-induced deformation and detachment phenomena, in: J. Ind. Microbiol. Biotechnol., J Ind Microbiol Biotechnol, 2002: pp. 361–367. doi:10.1038/sj.jim.7000282.
[30] S.C. Chew, B. Kundukad, T. Seviour, J.R.C. Van der Maarel, L. Yang, S.A. Rice, P. Doyle, S. Kjelleberg, Dynamic remodeling of microbial biofilms by functionally distinct exopolysaccharides, MBio. 5 (2014) 1–11. doi:10.1128/mBio.01536-14.
[31] G. Tierra, J.P. Pavissich, R. Nerenberg, Z. Xu, M.S. Alber, Multicomponent model of deformation and detachment of a biofilm under fluid flow, J. R. Soc. Interface. 12 (2015) 20150045. doi:10.1098/rsif.2015.0045.
[32] I. Klapper, C.J. Rupp, R. Cargo, B. Purvedorj, P. Stoodley, Viscoelastic fluid description of bacterial biofilm material properties, Biotechnol. Bioeng. 80 (2002) 289–296. doi:10.1002/bit.10376.
[33] J. Yan, C. Fei, S. Mao, A. Moreau, N.S. Wingreen, A. Košmrlj, H.A. Stone, B.L. Bassler, Mechanical instability and interfacial energy drive biofilm morphogenesis, Elife. 8 (2019). doi:10.7554/eLife.43920.
[34] M.J. Chen, Z. Zhang, T.R. Bott, Direct measurement of the adhesive strength of biofilms in pipes by micromanipulation, Biotechnol. Tech. 12 (1998) 875–880. doi:10.1023/A:1008805326385.
[35] H.-C. Flemming, Microbial Biofouling: Unsolved Problems, Insufficient Approaches, and Possible Solutions, in: Springer, Berlin, Heidelberg, 2011: pp. 81–109. doi:10.1007/978-3-642-19940-0_5.
[36] M.J. Chen, Z. Zhang, T.R. Bott, Effects of operating conditions on the adhesive strength of Pseudomonas fluorescens biofilms in tubes, Colloids Surfaces B Biointerfaces. 43 (2005) 61–71. doi:10.1016/j.colsurfb.2005.04.004.
[37] E.P. DeBenedictis, J. Liu, S. Keten, Adhesion mechanisms of curli subunit CsgA to abiotic surfaces, Sci. Adv. 2 (2016) e1600998. doi:10.1126/sciadv.1600998.
[38] A. Wang, S. Keten, Adhesive behavior and detachment mechanisms of bacterial amyloid nanofibers, Npj Comput. Mater. 5 (2019) 1–8. doi:10.1038/s41524-019-0154-7.
[39] H. Huang, G. Ye, C. Qian, E. Schlangen, Self-healing in cementitious materials: Materials, methods and service conditions, Mater. Des. 92 (2016) 499–511. doi:10.1016/j.matdes.2015.12.091.
[40] H.J. Chen, C.F. Peng, C.W. Tang, Y.T. Chen, Self-healing concrete by biological substrate, Materials (Basel). 12 (2019) 1–16. doi:10.3390/ma1224099.
[41] N. De Belie, E. Gruyaert, A. Al-Tabbaa, P. Antonaci, C. Baera, D. Bajare, A. Darquennes, R. Davies, L. Ferrara, T. Jefferson, C. Litina, B. Miljevic, A. Otlewska, J. Ranogajec, M. Roig-Flores, K. Paine, P. Lukowski, P. Serna, J.M. Tulliani, S. Vucetic, J. Wang, H.M. Jonkers, A Review of Self-Healing Concrete for Damage Management of Structures, Adv. Mater. Interfaces. 5 (2018) 1–28. doi:10.1002/admi.201800074.
[42] M. Seifan, A. Berenjian, Microbially induced calcium carbonate precipitation: a widespread phenomenon in the biological world, Appl. Microbiol. Biotechnol. 103 (2019) 4693–4708. doi:10.1007/s00253-019-09861-5.
[43] C.M. Heveran, S.L. Williams, J. Qiu, J. Artier, M.H. Hubler, S.M. Cook, J.C. Cameron, W. V Srubar III, Biomineralization and Successive Regeneration of Engineered Living Building Materials, Matter. 2 (2020) 481–494. doi:10.1016/j.matt.2019.11.016.
[44] Z.B. Bundur, S. Bae, M.J. Kirisits, R.D. Ferron, Biomineralization in self-healing cement-based materials: Investigating the temporal evolution of microbial metabolic state and material porosity, J. Mater. Civ. Eng. 29 (2017) 1–8. doi:10.1061/(ASCE)MT.1943-5533.0001838.
[45] A.M. Duraj-Thatte, N.-M.D. Courchesne, P. Praveschotinunt, J. Rutledge, Y. Lee, J.M. Karp, N.S. Joshi, Genetically Programmable Self-Regenerating Bacterial Hydrogels, Adv. Mater. 31 (2019) 1901826. doi:10.1002/adma.201901826.
[46] M. Schaffner, P.A. Rühs, F. Coulter, S. Kilcher, A.R. Studart, 3D printing of bacteria into functional complex materials, Sci. Adv. 3 (2017) eaao6804. doi:10.1126/sciadv.aao6804.
[47] S. Balasubramanian, M.E. Aubin-Tam, A.S. Meyer, 3D Printing for the Fabrication of Biofilm-Based Functional Living Materials, ACS Synth. Biol. 8 (2019) 1564–1567. doi:10.1021/acssynbio.9b00192.
[48] B. Pabst, B. Pitts, E. Lauchnor, P.S. Stewart, Gel-entrapped Staphylococcus aureus bacteria as models of biofilm infection exhibit growth in dense aggregates, oxygen limitation, antibiotic tolerance, and heterogeneous gene expression, Antimicrob. Agents Chemother. 60 (2016) 6294–6301. doi:10.1128/AAC.01336-16.
[49] N. Kandemir, W. Vollmer, N.S. Jakubovics, J. Chen, Mechanical interactions between bacteria and hydrogels, Sci. Rep. 8 (2018) 1–11. doi:10.1038/s41598-018-29269-x.
[50] J. Condi Mainardi, K. Rezwan, M. Maas, Embedding live bacteria in porous hydrogel/ceramic nanocomposites for bioprocessing applications, Bioprocess Biosyst. Eng. 42 (2019) 1215–1224. doi:10.1007/s00449-019-02119-4.
[51] D.T. Schmieden, S.J. Basalo Vázquez, H. Sangüesa, M. Van Der Does, T. Idema, A.S. Meyer, Printing of Patterned, Engineered E. coli Biofilms with a Low-Cost 3D Printer, ACS Synth. Biol. 7 (2018) 1328–1337. doi:10.1021/acssynbio.7b00424.
| Attachment | Size |
|---|---|
| Picture 1.png | 433.35 KB |
| Picture 2.png | 57.5 KB |
| Picture 3.png | 691.56 KB |
| Picture 4.png | 428.8 KB |
{kind=link}
{kind=link}
{kind=link}
{kind=link}